When discussing the topic of mechanism, two primary questions immediately leap to mind: "How do they find their way home?" and "How does a small fish make such a big journey?" The hormonal cascade that occurs immediately prior to and during migration to fulfill the physiological and metabolic needs of such a migration is an interesting avenue of research. Unfortunately, our understanding of the full developmental implications of these mechanisms in wild salmon is still very much incomplete. Due to the differences in behavior and physiology between hatchery-raised and wild salmon, it is important to remember that these findings only give a partial snapshot, and not a completed picture.
|
|
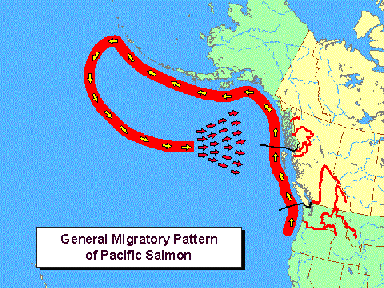
Left: Map of the North American coast of the Pacific Ocean displaying a generic migration route.
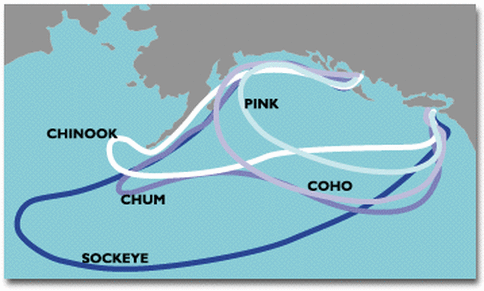
Right: Map of the Canadian and Alaskan coasts with ranges mapped to their respective salmon species.
Right: Map of the Canadian and Alaskan coasts with ranges mapped to their respective salmon species.
Pathfinding: Magnetoreception
Once the salmon has finished its time at sea, it must journey all the way from the ocean back to its home stream in order to mate, spawn, and die. It is very typical for this trip to be several hundred miles in either direction. Because of this, it is necessary for the salmon to navigate in manners both approximate and precise.
In the first stage of return from ocean to river, it is believed that salmon navigate via magnetoreception. Magnetoreception is the broad name for the family of sensory modalities that facilitate orientation based on the Earth's magnetic field, a trait which has evolved independently in many invertebrate and vertebrate animals. While the precise physiological mechanisms are still debated, there is a strong argument for the role of biogenic magnetite in these modalities. Magnetite believed to be endogenous has been found in salmon, as well as honeybees, birds, and sea turtles [5]. Ontogenic studies suggest that in Sockeye salmon, the presence and relevance of magnetite increases as the fish ages, indicating a greater role of magnetoreception in adult relative to juvenile pathfinding [6].
In the first stage of return from ocean to river, it is believed that salmon navigate via magnetoreception. Magnetoreception is the broad name for the family of sensory modalities that facilitate orientation based on the Earth's magnetic field, a trait which has evolved independently in many invertebrate and vertebrate animals. While the precise physiological mechanisms are still debated, there is a strong argument for the role of biogenic magnetite in these modalities. Magnetite believed to be endogenous has been found in salmon, as well as honeybees, birds, and sea turtles [5]. Ontogenic studies suggest that in Sockeye salmon, the presence and relevance of magnetite increases as the fish ages, indicating a greater role of magnetoreception in adult relative to juvenile pathfinding [6].
Pathfinding: Chemosensation
As salmon leave the ocean and return to their natal rivers, chemical senses take over the rest of the responsibility for getting the salmon back home. The traditional model of chemosensation describes a process of deep learning. At developmentally critical junctures, salmon will "imprint" on certain scents as indicators of their natal rivers. Later in life, these learned associations allow the salmon to discriminate which route is being fed by their birth river. Classic experiments testing this hypothesis used artificial odorants as a means of distinguishing "birth paths" - fish exposed to these articifial odorants early in life were then able to use that odor as a means of pathfinding to successfully return to their site of rearing [7]. Later experimentation developed and supported this hypothesis, with later studies indicating a sensitivity to the levels of other biologically relevant small molecules like amino acids.
One study suggests that in Pink Salmon, one of the most widely distributed species of salmon, imperfect homing accuracy can inform preference, choices, and behavior without producing a deterministic result. This 2011 study by Hiroshi Ueda implies a complex role of this chemosensory palate in route determination. In this study, it was found that altering the levels of one characteristic molecules in a water sample may not be sufficient to induce a differential response to otherwise similar water. When given a choice between natal stream water and natal stream water with a characteristic molecule removed (the amino acid glutamic acid), there was no significant difference in water preference. However, in comparison to dissimilar water samples, there was a preference for the modified natal water which was almost (but not quite) as strong as the preference for natal water. Implied by these results is a complex choice system not singly determined by the presence or absence of single identifiers, but a hierarchy of preference in which natal water may be most preferred, but other similar routes may also be favorable. The authors speculate there may be an evolutionary advantage for this imperfect selectivity - if the characteristics of the water influence a salmon's preference for a route but doesn't absolutely determine it, then a population of salmon could return selectively to the favorable conditions of the natal river that spawned them and also reap the benefits of a wide distribution where they generalize their pathfinding to find other suitable habitats. [8]
One study suggests that in Pink Salmon, one of the most widely distributed species of salmon, imperfect homing accuracy can inform preference, choices, and behavior without producing a deterministic result. This 2011 study by Hiroshi Ueda implies a complex role of this chemosensory palate in route determination. In this study, it was found that altering the levels of one characteristic molecules in a water sample may not be sufficient to induce a differential response to otherwise similar water. When given a choice between natal stream water and natal stream water with a characteristic molecule removed (the amino acid glutamic acid), there was no significant difference in water preference. However, in comparison to dissimilar water samples, there was a preference for the modified natal water which was almost (but not quite) as strong as the preference for natal water. Implied by these results is a complex choice system not singly determined by the presence or absence of single identifiers, but a hierarchy of preference in which natal water may be most preferred, but other similar routes may also be favorable. The authors speculate there may be an evolutionary advantage for this imperfect selectivity - if the characteristics of the water influence a salmon's preference for a route but doesn't absolutely determine it, then a population of salmon could return selectively to the favorable conditions of the natal river that spawned them and also reap the benefits of a wide distribution where they generalize their pathfinding to find other suitable habitats. [8]
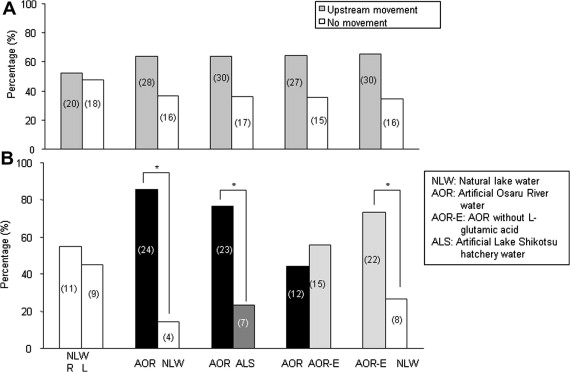
Above: Results from Ueda's study showcasing selective response to different water types. [8]
Hormone Regulation
Additionally, stages of salmon migration appear to be multiply influenced and regulated by hormonal signals and changes. Older research indicated a regular correlation between thyroxine levels and developmental stages assocated with olfactory imprinting [7]. Newer research suggests that projections from the olfactory system may cause stimulation of Gonadotropin-releasing hormone, in turn causing hormonal cascades that prepare the salmon for breeding as they approach their natal river to spawn [8].
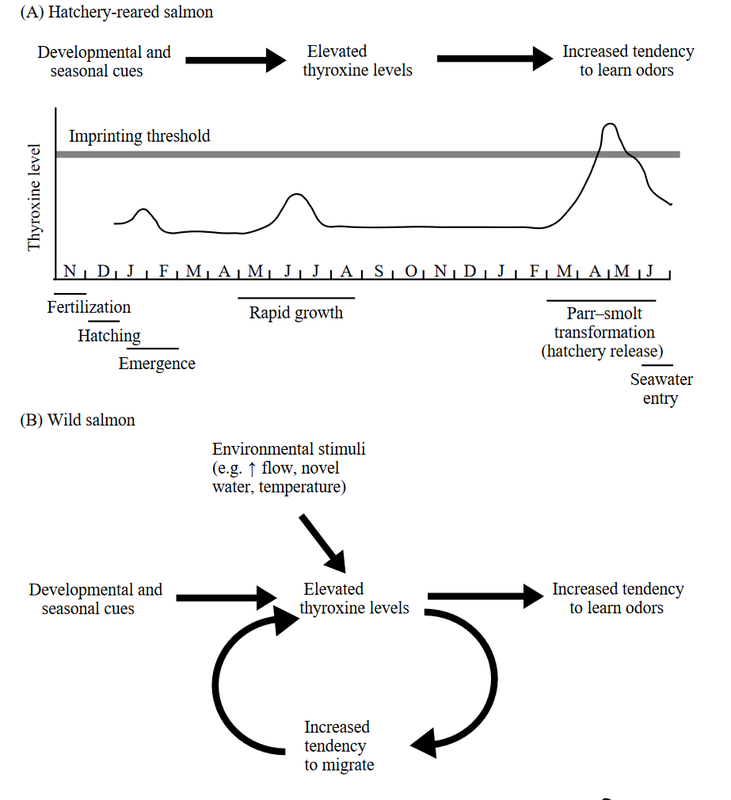
Above: A figure from the foundational Walker et al. paper [7] describing hormonal fluctuations in hatchery salmon and speculating how these hormonal mechanisms may describe the ontogeny of wild salmon.
